Що говорить ДНК-генеалогія про кавказців

Але цей нарис - не про духовну близькість , а про ту, що записана в наших ДНК. І ось тут з'ясовується, що ми з багатьма кавказцями брати вельми далекі. Втім, це ясно і при погляді на антропологію, і на написання знаків писемності, і на звучання мов. І ми, і кавказці - європеоїди, але якщо росіяни з антропології в переважній більшості кавкасіонці, то кавказці по відомим (і завжди спірним) класифікаціями розходяться на кавкасіонську, вірменоїдну, понтійську і каспійську антропологію. Тут, до речі, термін «кавказької», або «кавкасоідна» (раса) зовсім не означає «кавказька», і сама ця назва має цікаву історію. У 1795 році німецький антрополог Йоганн Фрідріх Блуменбах знайшов на Кавказі череп, який видався йому верхом досконалості і витонченості пропорцій. Природно, він належав європеоїдам, такий гарний, кому ж ще?
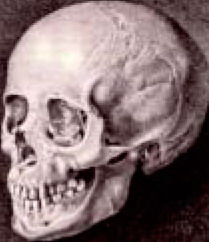
Ось той самий «красивий череп», який і поклав початок терміну «кавкасіонська раса». Блуменбах вважав, що біла людина, або европеоїд, з'явився спочатку на Кавказі, і тому племена, що жили на Кавказі, він вважав зразком найбільш чистого і незмішаної типу «білої раси». Так і прийнялося в науковій літературі, а потім і в побуті, як «кавкасіон» для позначення людей європейського походження, хоча те, що європеоїди виникли на Кавказі, так і залишилося недоведеним. Де виникли європеоїди - не ясно. Може, і на Кавказі. Може, в Центральній Європі. Може, на Руській рівнині. Надійних даних в науці поки немає.
Так от, до кавказької антропології на Кавказі відносяться, наприклад, гірські грузини - свани, хевсури, а також осетини, черкеси, лезгини, карачаївці, аварці, інгуші, чеченці, даргинці, лакці, кабардинці - загалом, перераховувати доведеться довго, на Кавказі більше 50 народностей. Арменоідна антропологія - у вірмен, ассірійців і східних грузин. Понтійська - у абхазів, адигів і західних грузин. Каспійська - у азербайджанців.
За мовам кавказці теж різняться - у них і індоєвропейські мови, і кавказькі, і тюркські. На індоєвропейських мовах говорять осетини, вірмени, курди. На кавказьких - грузини (картвельські мови), абхази і адиги (абхазо - адигські мови), інгуші і чеченці (нахскі мови), аварці, даргинці, лезгини, лакці (дагестанські мови). На тюркських - азербайджанці, балкарці, карачаївці, кумики, ногайці, турки-месхетинці.
Ясно, що походження кавказьких народів і народностей має бути різним, з різною історією, з різними шляхами міграцій. Що на цей рахунок говорить археологія, і як з цим узгоджуються дані ДНК-генеалогії?
Повинен зробити нелегку заяву. Є очевидні правила написання статей - або популярні, або для фахівців. Вони в цих двох випадках пишуться зовсім по-різному. Але це тоді, коли спочатку пише фахівець для фахівців, матеріал встановлюється, обкатується у фахових виданнях, а вже потім, коли-небудь, він спрощується до популярного викладу. Кому треба розібратися в деталях - будь ласка, є професійні статті. Але ДНК-генеалогія розвивається настільки стрімко, а інтерес до її знахідкам настільки великий, що часом спочатку пишуться популярні статті, а вже потім - професійні. У всякому разі, у мене так зазвичай і виходить.
У даному нарисі я об'єднав обидва підходи. Це і є нелегка заява з мого боку. У першій частині йде популярний виклад, і потім воно поступово починає переходити в напівпрофесійне, і потім практично в професійне. На це я пішов тому, що читачі на Переформат різні: одним треба просто ознайомитися з матеріалом, інші вимагають посилань на джерела, пояснень, як велися розрахунки, як будувалися дерева гаплотипів, обурюються, якщо автор не дає похибки розрахунків у формі стандартних квадратичних відхилень, а це в ДНК-генеалогії важливо. Тому я спробував написати матеріал так, щоб задовольнити вимоги і тих, і інших. Якщо розрахунки нецікаві, їх можна пропускати. Якщо похибки розрахунків неважливі, на них можна не звертати уваги. Якщо спадковий гаплотип нецікавий і занадто детальний, не звертайте на них уваги. Це ж стосується гілок гаплотипів і розрахунків по ним.
Але думаю, що якщо матеріал читатимуть кавказці, а вони читати неодмінно будуть, то вони стануть заглиблюватися в будь-які деталі, і стануть вимагати ще. Так що приношу всім вибачення - просто цікавим за зайві деталі викладу, професіоналам - за недостатні деталі викладу, кавказцям - за те, що так мало помістив деталей про кавказькі етноси і їх історії. В принципі, у мене є матеріал по ДНК-генеалогії Кавказу на сотні сторінок, і це притому, що число кавказьких гаплотипів дуже недостатньо для створення більш - менш адекватної картини родів і племен Кавказу з точки зору ДНК-генеалогії. Все це поступово буде публікуватися, у тому числі в співдружності з археологами, істориками, лінгвістами. Так що прийміть це як перше, попередню спробу доторкнутися до ДНК-генеалогії Кавказу.
Коротко - про археологію Кавказу. На Кавказі є безліч археологічних пам'яток, але ми коротко зупинимося тільки на найдавніших археологічних культурах, і тому зазвичай найбільш дискусійних. Саме на тих, де ДНК-генеалогія може привнести незалежні дані на користь тієї чи іншої версії походження археологічної культури. Нагадаю, що археологічні культури поділяються за сукупністю матеріальних ознак, які відносяться до однієї епохи і одній території, і по них, матеріальним ознаками, датуються - або радіовуглецевим методом, або «синхронізацією» з тими, що вже датовані.
Одна з найдавніших культур - це північнокавказька культура наколотою перловою кераміки кам'яного століття з переходом в ранній бронзовий вік (6500-5500 років тому), на території якої знаходяться сучасні Адигея і Карачаєво-Черкесія. Вона переходить в Майкопську культуру раннього бронзового століття (6000-5000 років тому, за іншими даними пізніше, 5200-4300 років тому), або, як її часто називають, майкопську культурну спільність, територія якої великою смугою проходить по передгір'ях Північного Кавказу від Чорного моря до сучасної Чечні, і сусідить на півночі з ямної культурою, на півдні - з куроаракскою культурою раннього бронзового століття (5500-4000 років тому). Остання займає частину Закавказзя і прилеглі території Близького Сходу, і є спадкоємицею культури ТМЗК (6500-5500 років тому), а та, в свою чергу, наступницею культури Сіоні (7000-6000 років тому).
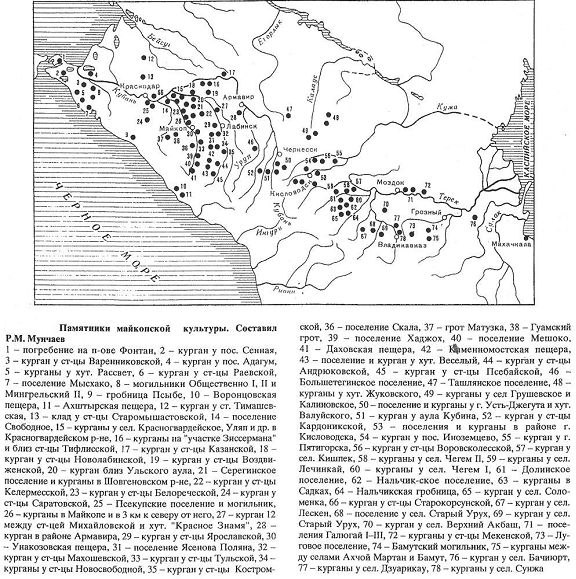
Майкопска культура переходить в північнокавказьку культуру бронзового століття (приблизно 4200-3800 років тому).
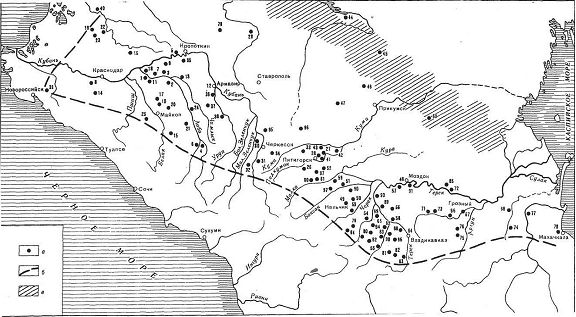
Ареал поширення північнокавказької культурно-історичної спільності (4200-3800 років тому), спадкоємиці майкопської культури (6000-5000 або 5200-4300 років тому, за різними оцінками). На карті показано географічне розташування археологічних пам'яток і умовна межа культурно-історичної спільності (пунктир), а також штрихуванням зона стику культур (по Бизову 2011).
На Заході Грузії і у Вірменії відома триалетська археологічна культура бронзового століття (4000-3500 років тому), що переходить в колхидску культуру бронзового і залізного століть (3600-2700 років тому). На Центральному та Західному Кавказі - кобанська культура (3300-2300 років тому), з її переходом від бронзового до залізного віку. Список археологічних культур Кавказу цим далеко не закінчується, і ці культури наведені тут для деяких історичних ілюстрацій.
Треба відзначити, що археологи звичайно датують культури за принципом не «років тому», а по «до нашої ери». Але оскільки ДНК-генеалогія оперує хронологією за принципом «років тому», то доводиться для стикування даних використовувати незвичну для археологів хронологічну систему.
Як звичайно, шляхи виникнення археологічних культур активно дебатуються. Як, втім, і все інше, пов'язане з археологічними культурами.
Нерідко буває, що у культур виявляються коріння, що розходяться в різні сторони. Мешканці культур часом змінювали один одного, причому ці були зовсім не родичі - це були зовсім різні роди, які прибули з різних далеких країв. У кавказьких культур таких мігрантів було, щонайменше, три важливих потока, які іноді йшли хвилями, протягом великих часів. Археологи в чому сходяться на тому, що найважливіший міграційний потік йшов на Кавказ 7000-6500 років тому і пізніше, прибуваючи з Месопотамії та Близького Сходу - Сирії, Східної Анатолії. Цей потік деякі пов'язують з прибуттям убейдо-урукских традицій і з семітськими мовами, але останнім часом його відносять до дещо пізнішої урукскої культури (приблизно 6400-6000 років тому). З півночі йшли мігранти степових культур, хвалинської культури Поволжя (7000-6600 років тому), азово-дніпровської культури (7200-6800 років тому) - це, по всій видимості, ербині, носії гаплогрупи R1b. Пізніше, приблизно 4600-4500 років тому, з півночі прибули засновники кобанської культури, і сама культура в тому вигляді, в якому ми її знаємо, склалася 3300-2300 років тому. Це, цілком можливо, носії гаплогрупи R1a, з їх індоєвропейськими мовами.
Дослідники залучають характеристики поховальних обрядів, що розглядається як одна з найбільш стійких особливостей древніх культур. Відзначається, що домайкопске населення укладала небіжчиків на спині з підігнутими ногами, головою на схід; ранні майкопці клали померлих у скорченому положенні - чоловіків на правому боці, жінок на лівому, головою на південь; пізніше укладали головою на захід або південний захід. Пізні майкопці, з переходом в північнокавказьку культуру, укладали небіжчиків на спину у витягнутому положенні. Це теж може бути вказівкою на різне коріння майкопської культури і на різні культури - спадкоємиці. Якщо залучити нечисленні дані ДНК-генеалогії, то складається враження, що домайкопске чоловіче населення було ербині, носіями гаплогрупи R1b, прийшли з півночі. Дійсно, в попередній частині згадуються мігранти північних степових культур 7200-6600 років тому, це і є домайкопскі часи. Пізні майкопці могли бути носіями гаплогрупи R1a, аріями, на їх міграційному шляху через Кавказ в Месопотамію, Сирію (мітаннійскі арії), Аравійський півострів.
Дійсно, було відзначено, що носії гаплогрупи R1a в Європі (нинішня Німеччина) укладали небіжчиків-чоловіків в скорченому положенні на правому боці (головою на захід), жінок на лівому (головою на схід), і всіх обличчям саме на південь (Haak et al, 2008), як і ранні майкопці. За часом підходить - арії йшли через Кавказ на південь приблизно 4500-4000 років тому (Klyosov and Rozhanskii, 2012). Хто укладав небіжчиків на спину у витягнутому положенні - поки неясно, це могли бути носії або гаплогрупи G2a, або J2. Тут треба збирати і систематизувати дані. До цього можна відзначити, що недавні розкопки на Чорноморському узбережжі сучасної Болгарії (район Варни), в місцях древнього видобутку солі, з датуваннями від 7500 до 6500-6200 років тому, показали трупоположення тих людей в скорченому положенні на правому боці (Current World Archaeology, August/September 2013, vol. 5, No. 12, 24-31), що знову може вказувати на носіїв гаплогрупи R1a. Правда, недалеко було знайдено поховання, в якому чоловік лежав у скорченому положенні на лівому боці. Або похоронні обряди насправді були нестійкими, що серйозно суперечить основній парадигмі археології, або друге поховання не мало відношення до першого, і могло ставитися до іншої епохи, іншому людському роду. Або, нарешті, було переплутано підлогу кістяка у похованні. З цитованої статті це неясно. Поруч - ще одна потрійна могила, двоє чоловіків і одна жінка - всі троє в скорченому положенні на правому боці, головою на північ. Правда, обидва чоловіки вбиті, черепа пробиті гострим предметом. Можливо, чужаки, і тому підлягали іншому обряду поховання?
Відносно майкопської культурної спільності було чимало гіпотез щодо шляхів її виникнення - наприклад, передбачалося, що мігранти з півдня просувалися на північ уздовж Чорноморського узбережжя, і вже значно пізніше поширилися на схід. Однак радіовуглецевий аналіз східної групи майкопських пам'яток (мітка 71 у Терека, на карті вище) дав датування 4540±70 років тому, 4660±80 років тому, 4990±120 років тому (Кореневский, 1993, цит. по Бызов, 2011), що, на думку деяких дослідників, показує синхронність західного і східного варіантів майкопської культури. Як пише І. Бизов (2011), «як би там не було, але питання про шляхи міграції майкопців з Передньої Азії на Північний Кавказ залишається поки відкритим»
Найбільш інтригуючим є, мабуть, прибуття на Кавказ урукскої культури між 7 і 6 тисяч років тому, представники якої розселилися від південного до північного Кавказу, послідовно заснувавши важливу археологічну культуру Тсопі (яка також продовжила традиції культури Сіоні), куро-аракську культуру, майкопську культуру, і принісши з собою характерні традиції Урука, стародавнього міста-держави, що виникла на території між сучасними Багдадом і Басрой в Іраку не пізніше 7 тисяч років тому. Зараз на місці Урука знаходиться місце під назвою Варка, і воно практично нежиле. Річка Євфрат давно пішла з цих місць, а колись, близько п'яти тисяч років тому, в Уруці на території в 6 квадратних кілометрів жили більше 50 тисяч чоловік (див. журнал Archaeology, вересень-жовтень 2013 р., Стор. 26-32). Тоді це був найбільше місто на Землі, його пізній суперник - Вавилон - виник через дві тисячі років. Урук як місто простояв п'ять тисяч років, поки не був залишений через відхід Євфрату і відповідно зміненого клімату, комерційної конкуренції, зміни політичної карти тих країв.
Питання - чому в ті давні часи представники культури Урука рушили на Кавказ, притому, як виявилося, щоб не повертатися назад, а оселитися назавжди? Знаючи красу Кавказу, останнє не дивно. Але що привело туди людей з Близького Сходу? Відповідь дають археологи - урукці прийшли на Кавказ за цінними металевими рудами - .мідь, срібло, золото, а також, мабуть, йдучи від перенаселення (Pitskhelauri, 2012, Bulletin of the Georgian Academy of Science, т. 6, № 2, 153-161)
Це - дуже стислі уявлення сучасних археологічних наук. Пізніше в цьому нарисі ми приміряємо на ці уявлення результати і висновки ДНК-генеалогії
Спільним є положення, що на Кавказі - безліч етнічних груп, і в кожній долині і на кожному нагір'ї свої народності, свої мови або діалекти. Це, загалом, вірно, але ДНК-генеалогія, яка займається походженням родів і племен, несподівано показала, що родів на Кавказі не так багато, всього декілька. Вони перемішані в різних пропорціях у різних етнічних групах, що і дало різноманітність, мозаїку ДНК-генеалогічних ліній. Але є відмінності і більш високого порядку, і вони ділять Кавказ практично навпіл. Одна половина - західна, в основному гаплогруппа G2a, інша половина - східна, в основному гаплогрупи J2 і J1, кожна з абсолютно різною історією. І в усі прошарком, але різною мірою, часом сходить нанівець, входять гаплогрупи R1a і R1b, кожна знову абсолютно зі своєю, різною історією.
Спробуємо в цьому розібратися. Але для цього розглянемо спочатку, що таке по суті ДНК-генеалогія..
Основні положення ДНК-генеалогії. У даному нарисі йтиметься про нову науку, яка тільки створюється, основи якої тільки закладаються в останні роки. Якщо точніше, то основи цієї нової науки вже створені, причому стрімко, і йде прогресивне накопичення експериментального матеріалу. Новий матеріал надходить потоком, щодня в бази даних йдуть десятки і сотні нових «експериментальних точок», які за принципом зворотного зв'язку коригують методологію нової науки, що призводить до уточнення методів розрахунку.
Ім'я цієї науки - ДНК-генеалогія. Її експериментальні дані - це картина мутацій в нерекомбінантних ділянках чоловічої статевої хромосоми (насправді - і в мітохондріальної ДНК, але в цьому нарисі мова піде тільки про Y-хромосому, інакше обсяг його вийде за межі допустимого), причому картина мутацій як в Y-хромосомах окремих людей, так і їх груп, популяцій. Методологія нової науки - переклад динамічної картини мутацій в хронологічні показники, в часи життя спільних пращурів популяцій, а насправді - спільних пращурів давніх родів і племен. Тобто фактично проводиться розрахунок часів, коли в давнину жили ці роди і племена.
Мутації, що розглядаються в ДНК-генеалогії - це або поодинокі (як правило) заміни нуклеотидів в ДНК, як, наприклад, аденін на цитозин, або цитозин на тимін, або вставки нуклеотидів, або делеції, або мутації складніші, при яких помилка копіює ферменту призводить до переносу цілого блоку нуклеотидів, тандемного, як його часом називають. Це дає або подовження серії таких блоків на один (рідко - відразу на два-три блоки), або їх укорочення. Перші мутації - SNP (Single Поліморфізм, або в перекладі «поодинокі нуклеотидні варіації»), або «БНіП», зазвичай дуже стабільні, і їх для ДНК-генеалогії відбирають тільки такі, які трапляються лише один раз (два рази - максимум) за історію людства. Тому вони є маркерами родів людства. Приклади будуть дані нижче.
Мутації другого типу - STR (Short тандемних повторів, або «короткі тандемні повтори") - значно більш швидкі, і відбуваються в певних ділянках, або локусах, або маркерах (це все синоніми) ДНК раз на кілька десятків або сотень поколінь. Набір цих маркерів складає гаплотип, приклади гаплотипів будуть дані нижче. Гаплотип - це по суті ланцюжок чисел, що показують число повторів у певних маркерах. Тому гаплотипи ДНК вибирають так, щоб у них таких маркерів було якомога більше (але все-таки щоб залишатися в рамках практичності), і в ранніх роботах використовувалися 6-маркерні гаплотипи, потім 12-маркерні, 17- і 19-маркерні, потім 25- і 37-маркерні, а зараз робота рутинно ведеться з 67- і 111-маркерними гаплотипами (правда, в академічних публікаціях це звичайно від 8 до 17 маркерних гаплотипів). У 67-маркерних гаплотипах, наприклад, одна мутація відбувається в середньому за 8 поколінь, в 111-маркерних - за 5 поколінь.
Оскільки вже показано і доведено, що ці мутації в маркерах відбуваються в основному (або виключно) неупорядкованно, то до них виявилося можливим застосовувати правила і підходи хімічної кінетики, або біологічної кінетики, кому який термін більше подобається - суть одна. І БНіП, і маркери мутують за законами кінетики першого порядку, інших закономірностей поки не виявлено. Це означає, що кожному маркеру властива певна константа швидкості мутації, і кожному гаплотипу (що складається з набору маркерів) - теж властива певна середня константа швидкості мутації, що дорівнює сумі констант швидкостей мутацій окремих маркерів. Тому чим древній загальний пращур популяції, тим більше мутацій накопичується в гаплотипі його нащадків в порівнянні з ним, тим більше поколінь (і років) відокремлює сучасних нащадків від їх загального прашура, і це число поколінь (і років) розраховується методами ДНК-генеалогії. Як це робиться - теж буде показано нижче на багатьох прикладах.
ДНК-генеалогія базується на чітких правилах, яких можна нарахувати тринадцять.
Перше - до генетики ДНК-генеалогія прямого відношення не має. Хоча в певній частині ДНК-генеалогії розглядають і гени, наприклад, при розгляді всього генома людини, але це - спеціальна частина ДНК-генеалогії, і цього тут не буде. ДНК - це не тільки гени. Гени взагалі займають тільки 1,9 % послідовності ДНК. Решта 98,1 % - довгі ділянки, так звані «нікчемні». Генів там немає (це детально розібрано в статті - Кльосов, 2012).
У ДНК є протяжні послідовності повторюваних нуклеотидів, в яких, повторюю, генів немає, і ці послідовності детально копіюються при передачі спадкової інформації від батька до сина. Я спеціально пишу «від батька до сина», тому що в даному нарисі йдеться тільки про «чоловічий» спадкової інформації, переданої з чоловічої статевої Y-хромосомою. У жінок її, цієї хромосоми, немає.
«Нікчемними» ці негенні послідовності ДНК назвали через незнання, і ця назва вже відходить у минуле. У міру досліджень, виявилося, що ці послідовності зберігають масу інформації і не тільки «генеалогічного» характеру. Більше того, виявилося, що вони, ці «нікчемні» ділянки, містять сотні, якщо не тисячі крихітних генів РНК, які кодують так звані мікро-РНК, довжиною всього пару десятків нуклеотидів кожен, і які регулюють синтез білків. Але це - предмет іншої розповіді.
Ці негенні ділянки ДНК в чоловічій Y-хромосомі, однієї з 23 хромосом, які доставляються сперматозоїдом в організм майбутньої матері, копіюються від батька до сина, покоління за поколінням. Оскільки у матері такої хромосоми немає, то син успадковує її тільки від батька. Хромосоми сплітаються з материнськоою ДНК і утворюють нову комбіновану молекулу ДНК, в якій хромосоми батька і матері перетасовуються. Перетасовуються всі, крім чоловічої Y-хромосоми.
Так і виходить, що батько передає синові цю Y-хромосому інтактною, суворо скопійованої зі своєї. А свою Y-хромосому він отримав від свого батька. Той - від свого. І так далі, на тисячі, десятки тисяч і сотні тисяч років углиб, назад, до далеких пращурів сучасних людей і до пращурів тих далеких пращурів.
Тут - важлива примітка. В антропології прийнято вважати, згідно з базовими поняттями, парадигмою антропології, що сотні тисяч років тому людей не було. Були палеонтропи, архантропи, неандертальці, еректуси, інші гомініди. Однак наші предки були, як їх не називати. Вони були і десятки, і сотні тисяч років тому, і мільйони років тому. Від них, «приматів», ми й успадковуємо гаплотипи і гаплогрупи, нехай значною мірою змінені, мутованих, іноді втративши цілі шматки.
Таким чином, аргументи, що тоді-то давно не було пращурів сучасної людини, в принципі невірні. Вони були, але просто або не виявлені антропологами (що цілком можливо), хоча і були на розглянутій території, або вони були представлені мінорною долею гомінідів та інших «приматів», і при вивченні останніх просто не зустрічалися, проскочили сито антропологів, вельми крупноячистої. Або визначення, що використовується антропологами, занадто жорстке і не відображає значно складнішу історію походження людства.
Якби Y-хромосома так і передавалася з покоління в покоління дійсно незмінною, толку для генеалогії від неї було б мало. Але незмінного нічого в світі немає, особливо коли мова про копіювання. Копіювання без помилок не буває. У тому числі і копіювання Y-хромосоми.
Звідси друге положення - час від часу при копіюванні Y-хромосоми в копії проскакують помилки. Фермент під назвою ДНК-полімераза (насправді разом з цілим набором молекулярних інструментів) або просто помиляється і робить деякі ділянки ДНК коротше або довше, подовжуючи або скорочуючи повтори нуклеотидів, або «ремонтує» пошкоджені ділянки і в ході «ремонту» видаляє ушкодження (ця операція називається «делеція») або «вшивається» новий нуклеотид (називається «вставка» або «інсерт»). Те, що вдалося відремонтувати, в мутації не потрапляє, ніби мутації і не було. Що не вдалося - передається синові при народженні. Якщо народилася дочка, то Y-хромосома не передається. Якщо тільки дочки, або дітей немає - Y-хромосома термінується, разом з чоловічою спадковою лінією.
Мутації, які цікавлять ДНК-генеалогію, бувають двох типів - або зміна числа повторів, тандемів (STR), або «точкові», одно-або кілька - нуклеотидні (SNP).
Виявилося, що багато повторів нуклеотидів знаходяться у різних людей в одних і тих же ділянках Y-хромосоми. Ці ділянки вже фахівцями пронумеровані, класифіковані, зведені в списки. Вони отримали назву «маркери». Набір маркерів, точніше, повторів у них, називається «гаплотип». Вже відомих маркерів - більше тисячі. Вони, як правило, одні і ті ж у всіх людей на Землі. Вони відрізняються тільки числом повторів, що викликано тими самими помилками ферментів при копіюванні з покоління в покоління. Уважний розгляд маркерів і гаплотипів дозволило зробити висновок, що всі люди на Землі походять від одного пращура.
Отже, положення третє: всі люди - родичі, вони всі походять від одного пращура. Цей пращур виявився древнішим, ніж передбачалося раніше, але він був. Ще недавно вважалося, що він жив приблизно 70 тисяч років тому, потім 120 тисяч, а тепер він вже йде глибше 200 тисяч років тому, поступово наближаючись до загального пращура з неандертальцем 300-500 тисяч років тому. Приблизно такий час знадобилося, щоб отримати ту розбіжність гаплотипів по мутаціям, яке спостерігається у всіх проживаючих нині людей. Останнє поглиблення за часом сталося тому, що в Африці знайшли незвичайні ДНК-генеалогічні лінії, які відрізняються від всіх людей на Землі, в тому числі і від інших африканців, але гаплотипи хоч і віддалено, але схожі на ті, що є у всіх нас. Значить, загальний пращур був один і той же, тільки дуже давній.
Положення четверте - гаплотипи зображують у вигляді числа тандемів, або повторів, по кожному маркеру, обраному з десятків і сотень. В англомовній літературі, як уже говорилося, їх називають STR, або Short Tandem Repeats. Самий простий і короткий гаплотип з тих, які розглядає ДНК-генеалогія, складається з п'яти або шести маркерів. Наприклад, у грузин гаплогрупи G2a , яка у них основна за чисельністю і становить близько половини чоловічого населення, базовий (або предковий) 6-маркерний гаплотип, в тому запису, який зазвичай приводять в наукових публікаціях, особливо в застарілих, такий:
15-12-22-10-10-14
Він показує число повторів нуклеотидних ділянок в маркерах, які позначають відповідними індексами 19, 388, 390, 391, 392 і 393. У першому маркері (номер 19) було знайдено 15 повторів певної послідовності нуклеотидів. У другому - 12 повторів вже іншої послідовності і так далі. При передачі цього гаплотипу від батька синові з імовірністю приблизно одна сота, або один відсоток (тобто в середньому у одного на сто народжень) може відбутися мутація, наприклад, така:
15-12-21-10-10-14
А може статися й така -
16-12-22-11-10-14
або будь-яка інша, тільки одні мутації в одних маркерах відбуваються частіше, в інших - рідше. А в середньому мутація в таких 6-маркерних гаплотипах відбувається, як було знайдено, в середньому раз на сто народжень хлопчиків. Якщо перевести в часи, то константа швидкості мутації в 6-маркерних гаплотипах дорівнює 0.0088 мутацій на весь гаплотип за 25 років.
Звідси випливає положення п'яте - за кількістю мутацій в гаплотипі можна визначати вік гаплотипу, тобто час, що минув від загального пращура цього гаплотипу до його сьогоднішніх нащадків. Оскільки сини зберігають гаплотип батька, переданий у спадок, і мутації в цьому гаплотипі проскакують в середньому лише раз приблизно в 3100 років (6-маркерний гаплотип) або раз на 1300 років (12-маркерний гаплотип), або раз на 125 років (111-маркерний гаплотип), то навіть через 5000 років у нащадків збережеться 23% початкового 6-маркерного гаплотипу, без змін. Тобто в списку з 100 гаплотипів нащадків - 23 гаплотипу будуть такими ж, який був у пращура 5000 років тому. Це якщо розглядати 6-маркерні гаплотипи. При розгляді 12-маркерних гаплотипів ті ж 23% гаплотипів пращура збережуться через 88 поколінь, або 2200 років.
Так що і через тисячоліття можна визначити гаплотип пращура. І з його вигляду можна дізнатися, з яких країв пращурк прийшов, порівнявши вид гаплотипу з гаплотипами по територіях, і з доступними копалинами гаплотипів.
Положення шосте - гаплотипи не вказують на етноси, це зовсім різні поняття. Гаплотипи вказують на давні роди, племена, які набагато старші етносів. З тих давніх часів гаплотипи давно розійшлися по різних територіях, на яких потім, через тисячоліття сформувалися етноси. Наприклад, 6-маркерний спадковий гаплотип племені G2a1a1, основний у грузин (з небагатьма мутаціями, що набігли за тисячоліття)
15-12-22-10-10-14
є і у осетин (дігорців і іронців), і у шапсугів, і у черкесів (адигів), і у абхазів, з деякими варіаціями, тому що загальний пращур в кожному етносі жив у різний час. До одних цей рід прийшов недавно і бурхливо розмножився, як у осетин, у інших був зі стародавніх часів, у третіх рід майже знищили вороги, залишилося мало, і пропорції родів в етносі хитнулися в інший бік. Конкретні цифри будуть дані нижче, по етносам. Але всі вони, нащадки гаплогрупи G і племені G2a1a1, брати, один і той же рід, все від одного і того ж пращура - патріарха, хоча частка цього роду у всіх зараз різна. Якщо у грузин його 32% (ще 10% - племені G2a3b1, і ще кілька відсотків племені G2a*), то у осетин - дігорців 55%, у осетин - іронців 72%, у абхазів 27%, у черкесів 14%, у шапсугів всього 1%.
Виходячи з вищесказаного зрозуміло, що просто частка у відсотках сама по собі означає не дуже багато, це просто структура сучасної популяції, те, чим займаються популяційні генетики. У осетин цього роду (G2a1a1) більше, ніж у грузин, але він молодший, тобто прийшов до Осетії пізніше. У осетин три гілки цього роду, віком 1675±380 років, 1375±210 років і 450±175 років - наша ера або навіть середні століття. У грузинів ці гілки йдуть в минулу еру, із загальним пращуром більше трьох тисяч років тому. А взагалі на північно-західному та центральному Кавказі загальний пращур роду G2a1a1 жив 4875±500 років тому, а батьківського роду G2a - 11500 років тому.
Інше плем'я тієї ж гаплогрупи (G2a3b1), з предковим гаплотипом
15 12 23 10 11 14
(Варіації виділені жирним шрифтом) увійшло до складу сучасних грузин в кількості 10%, осетин - дігорців 5%, осетин - іронців 2%, абхазів 24%, черкесів 38%, шапсугів 86%. Як бачимо, у черкесів і шапсугів крен гаплогрупи по кількості в інший бік. Рід той же, а плем'я інше. Інший субклад гаплогрупи, в термінах ДНК-генеалогії. Це вже не G2a1a1 - P18, G2a3b1 - P303, вік племені трохи більше 5 тисяч років тому, в межах похибки розрахунків такий же, як і племені G2a1a1. Спільний пращур обох племен, як було зазначено, жив 11500 років тому. Позначення вище показують два індекса, один - порядковий індекс гаплогрупи, або субклада (це часто за змістом синоніми, але гаплогруппа зазвичай означає рід, а субклад - плем'я, частина роду, хоча це часто умовності, і в складі великих племен є малі, і там і там - субклад), другий індекс - номер сніпа по каталогу. Часто їх показують два разом, тандемом, щоб не плутатися.
Але це - північно-західний і центральний Кавказ. А східний Кавказ? На східному Кавказі цієї гаплогрупи G2a мало - там, в основному, зовсім інші роди, J2 і J1. Племені G2a1a1 - P18, яке домінує у грузин і осетин, на сході майже немає, тільки 6 % у чеченців, і то тільки в Дагестані та Інгушетії, а в самій Чечні цього племені взагалі практично немає. Є 10% у аварів, але племені G2a3b1 - P303, якого мало у грузин і осетин. Є 13% у лезгин, але зовсім іншого давнього племен , G2a*.
Оскільки у кожного племені і в кожному етносі, в кожному регіоні можна визначати вік племені і взагалі популяції в цілому, то можна визначати, коли і в якому напрямку йшли стародавні міграції.
Положення сьоме - всі розрахунки виробляються з похибками. Точні цифри отримані бути не можуть в принципі. Справа в тому, що мутації невпорядковані, тому ми маємо справу зі статистикою. Чим більше вибірка, тим точніше розрахунки. Похибка залежить від числа гаплотипів у вибірці, від протяжності гаплотипів, від того, наскільки точно визначена і вивірена, відкалібрована константа швидкості мутації, від того, наскільки древній загальний пращур. Особливо неточно порівнювати мутації в парах гаплотипів людей. На двох гаплотипах мутація - інша могла додатися буквально в попередньому або нинішньому поколінні. Це може відразу призвести до додатку - відібранню тисячі років «в одному поколінні». А у великій вибірці різниця щодо нівелюється статистикою. Є вибірки в тисячі гаплотипів - там розрахунки, звичайно, точніші.
Але в ДНК-генеалогії часто важлива не абсолютна точність, а концептуальний висновок. Наприклад, якщо хтось стверджує, що носії гаплогрупи R1b (які зараз становлять приблизно 60% чоловічого населення Західної та Центральної Європи), жили там, у Європі, ще 30 тисяч років тому (а такими твердженнями, без розрахунків, була заповнена академічна література з даними питань до недавнього часу), а розрахунки показали 4800 років тому, то тут справа не в точності, а в принциповому виведенні. Те ж саме по ДНК-генеалогії Кавказу - якщо дані показують, що Кавказ заселявся носіями гаплогрупи J2 з Месопотамії приблизно 7-6 тисяч років тому, причому заселявся різними племенами і по різних територіях, і вже відомо яким саме, і в який час, то це дає важливі дані історикам і археологам, які продовжують гарячково сперечатися з даних питань.
До цього веде положення восьме - гаплотипи неабиякою мірою (але не завжди) пов'язані з певними територіями.
Але як таке може бути? А от як. В давнину більшість людей пересувалися племенами, родами. Рід, за визначенням, це група людей, пов'язаних спорідненістю. Тобто гаплотипи у них однакові або близькі. Пам'ятайте - одна мутація в середньому відбувається за тисячоліття? Проходили тисячоліття, чисельність родів деколи скорочувалася до мінімуму («пляшкове горлечко» популяції), і якщо вижив мав деяку мутацію в гаплотипі, то його нащадки вже «стартували» з цією новою мутацією, копіюючи її поколіннями у своїх ДНК, а чоловіки - у своїй Y-хромосомі. Деякі люди покидали рід по своїй або чужій волі - полон, втеча, подорожі, військові походи, і вижили починали новий рід на новому місці. У підсумку карта світу з точки зору ДНК-генеалогії вийшла плямистою, і кожна пляма деколи має свій превалюючий гаплотип - гаплотип роду. Часто він і є гаплотип пращура, який розпочав рід на даній території.
Але є ще один тип мутації - точкові мутації, сніпи (калька з англійської SNP, що в приблизному перекладі й означає «одинична нуклеотидна мутація», або «одинична нуклеотидна варіація»). Вони - практично вічні. Раз з'явившись, вони вже не зникають. Теоретично, в тому ж нуклеотиді може відбутися інша мутація, змінивши перших. Але нуклеотидів - мільйони, і ймовірність такої події вкрай мала. Всього в хромосомах є багато мільйонів сніпів, з яких в ДНК-генеалогії застосовуються багато сотень, і кожен сніп відповідає гаплогруппі, тобто роду, або субкладу, тобто племені, хоча ці дефініції застосовують дуже вільно, як пояснено вище. Двадцяти найбільш великим гаплогрупам, кожна з яких охоплює сотні мільйонів людей, привласнили літерні позначення від А до Т, приблизно в хронологічному порядку появи відповідного роду на планеті. Або, принаймні, в тому порядку, як вчені вважають ці роди з'явилися. Хоча ревізій на цей рахунок належить ще багато. Тільки що пройшла чергова - в список додали дві гаплогрупи, А00 і А0, які нещодавно виявили в Африці, і які стоять абсолютно осібно навіть до інших африканцям, не кажучи про європейців або азіатів.
Отже, положення дев'яте - людей можна класифікувати за стародавніми родами не тільки (і не стільки) з вигляду гаплотипів, але і по наявності певних сніпів. Наприклад, носії гаплогрупп А і В зараз в основному живуть в Африці (але де ці гаплогрупи з'явилися, поки незрозуміло, схоже, що не в Африці, і туди мігрували); гаплогруппа С зустрічається особливо часто серед монголоїдів і жителів Австралії і Океанії; гаплогрупи G знаходять в стародавній Європі (копалини гаплотипи), на Кавказі, в Передній Азії; гаплогруппа J - початково Близький Схід і звідти перейшла Кавказ не пізніше 6-7 тисяч років тому; гаплогрупи J1 досить багато на Кавказі, а також серед арабів і євреїв на Близькому сході, гаплогрупи J2 багато на Кавказі, в Месопотамії і серед жителів Середземномор'я; гаплогруппа L з'явилася, схоже, в Передній Азії, і розійшлася в різні боки - її підгрупа (субклад) L1 - в Індії, L2 - у Грузії, L3 - на східному Кавказі. Гаплогрупи N мають багато жителів Китаю, Сибіру, півночі Росії, Прибалтики та частини Скандинавії, куди й прибули носії гаплогрупи N зі сходу - Уралу і до того Південного Сибіру тисячоліття тому).
Гаплогруппа I - можливо, з Російської рівнини не менше 40-45 тисяч років тому, і після майже повного вимирання або винищення приблизно 4500 років тому зараз майже виключно знаходиться в Європі, де відродилася відносно недавно, 3600-2300 років назад (для I1 і I2, відповідно); гаплогрупи R1a і R1b з'явилися в Південному Сибірі після довгої міграції їхніх пращурів, носіїв гаплогрупи К, з якої утворилася гаплогруппа Р і потім Q і R, напевно, з Європи або з Російської рівнини на схід, далеко за Урал. Звідти гаплогрупи R1a і R1b прийшли в Європу, причому різними міграційними шляхами. R1a стала основною гаплогрупою Східної Європи, R1b - західної та центральної Європи.
Дане положення зводиться до того, що у кожного чоловіка є сніпи з певного набору, за яким можна віднести носія до певного стародавнього роду. Часи появи сніпів, звичайно вживаних у ДНК-генеалогії, відносяться до діапазону від 25-40 тисяч років для «старих» сніпів до 10-15 тисяч років для «молодих», але зараз вже виявляють сніпи, що утворилися всього 1000-1500 років тому. Наприклад, чверть усього ірландського чоловічого населення мають сніп (R1b - M222), який утворився близько 1500 років тому. У континентальній Європі його практично немає.
Положення десяте - гаплогрупи не просто відповідають певним родам, але утворюють певну послідовність, сходи гаплогрупп, що показують їх ієрархію - послідовні, ступінчаті перехід від точки розбіжності африканських і неафриканських популяцій (приблизно 160 тисяч років тому) до самої недавньої гаплогрупи R, утворилася приблизно 30-40 тисяч років тому. Ці сходи називаються філогенетичним деревом гаплогрупп і їх сніпів. Всі гаплогрупи і субклади на дереві повинні включати сніпи «вищестоящих» гаплогрупп і субкладів. Тобто принцип «драбинки» повинен виконуватися. Наступність вузлових родів людства повинна дотримуватися.
Все це робить філогенетичне дерево гаплогруп досить міцною і обгрунтованою структурою. У нього є, втім, слабке місце - його філогенія не показує, на якому континенті зародилося людство, звідки пішли гаплогрупи, починаючи з перших, на загальному стовбурі, що йде від нашого спільного пращура з сучасним шимпанзе. Говорячи мовою філогенії, дерево гаплогруп не «вкорінене». Укорінення дерева - результат інтерпретацій спостережень і доступних експериментальних даних.
Положення одинадцяте - в ДНК -генеалогії зазвичай оперують поколіннями. Покоління в контексті ДНК-генеалогії - це подія, яка відбувається чотири рази на століття. Чисельно і за часом воно близьке до тривалості покоління в життєвому сенсі цього слова, але не обов'язково дорівнює йому. Хоча б тому, що тривалість покоління не може бути точно або хоча б у середньому визначена, вона «плаває» в реальному житті в залежності від багатьох факторів, включаючи культурні, релігійні та побутові традиції, приблизно і в середньому від 18 до 36 років, хоча межі цього не визначені.
В давнину цей діапазон був, мабуть, помітно зміщений до першої величини. Тому використовувати настільки «плаваючу» величину для розрахунків в широких часових діапазонах і для різних народів не представляється можливим або розумним. Швидкості мутацій нами відкалібровані під умовно узяте покоління тривалістю 25 років. Якщо комусь більше подобається 30 років на покоління або будь-яку іншу кількість років, швидкості можна перекалібрувати, і в підсумку виявляться рівно ті ж величини в роках. Так що скільки років припадає на покоління - в даному випадку не має значення, тому що при розрахунках меншому числу років на покоління буде просто відповідати пропорційно більше число поколінь, і підсумкова величина в роках не зміниться.
Положення дванадцяте - тільки ті мутації в гаплотипі є сенс розраховувати, екстраполюючи до часу загального пращура, які підпорядковуються певним кількісним закономірностям. Іншими словами, ДНК-генеалогія оперує трьома експериментальними чинниками: 1) наявністю сніпів, що відносять людину до певного роду; 2) наявністю мутацій, що дозволяють оцінювати час що минув від загального пращура сукупності гаплотипов і - при великих вибірках - від початку самого роду, від самого далекого з предків нині живих нащадків даного роду (тобто тут вважаються самі мутації); 3) закономірності переходів гаплотипів в їх мутовані форми, без чисельного обліку самих мутацій (тобто тут мутації не враховуються, ввраховуються немутаційні гаплотипи).
Це дозволяє оцінити, наскільки достовірні розрахунки пращура по мутаціям і дає ще один, незалежний спосіб розрахунків. Методи, в яких враховуються мутації, називаються лінійним, квадратичним і пермутаційним, з яких найбільш простий лінійний метод. Метод, в якому враховуються немутаційні гаплотипи, називається логарифмічним. В принципі, він використовує формули хімічної кінетики першого порядку. У лінійному методі загальне число мутацій в серії гаплотипів ділиться на число гаплотипів і на константу швидкості мутації для даних гаплотипів. У логарифмічному методі береться логарифм відношення загального числа гаплотипів до числа немутагованих гаплотипів, і ділиться на константу швидкості мутації.
Положення тринадцяте - у більшості випадків результати розрахунків майже не залежать від розміру вибірки (при числі гаплотипів більше двох-трьох десятків), тобто вони стійкі до статистичних варіацій. Розмір вибірки збільшує точність, і то тільки до певних меж. Це, правда, відноситься до досить великим популяціям, які перемішалися за тисячоліття, але саме з такими зазвичай і працюють.
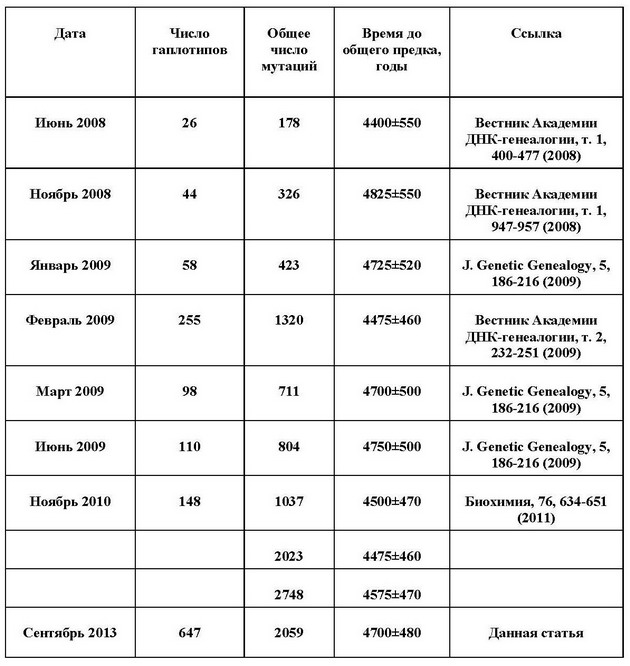
Наведемо приклад. У таблиці нижче наведені розрахунки часу до загального пращура гаплогрупи R1a на пострадянському просторі. Це, в основному, - Росія, Україна, Білорусія, Литва, Латвія, Естонія і Казахстан. Видно, що при збільшенні вибірки від 26 гаплотипів до більше шестисот результати практично однакові в межах похибки розрахунків.
на «пост-радянському просторі» по різних серіях гаплотипів
На малюнку приведено дерево реальної вибірки з 68 гаплотипів. Гаплотипи простенькі, 6-маркерні, загальний пращур всіх жив всього 625 років тому, так що мутацій в гаплотипі набігло мало. Можна навіть заздалегідь порахувати, скільки. Константа швидкості мутації для 6-маркерних гаплотипів дорівнює 0.0088 мутацій на гаплотип на умовне покоління в 25 років. Тоді за 625 років (25 умовних поколінь) у кожному гаплотипі набіжить 0.0088х25 = 0,22 мутації, і на всі 68 гаплотипів буде приблизно 0.22х68 = 15 мутацій. Приблизно - тому що дробові числа ми округляємо, не може бути «два землекопа і дві третини», якщо хто пам'ятає дитячі вірші С.Я. Маршака.
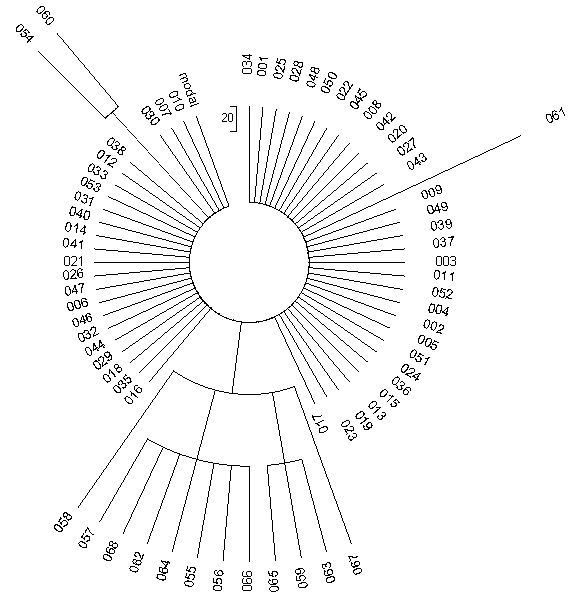
Наприклад, такі: 15 12 25 11 11 13. Вони і утворюють «гребінку» на дереві.
Дивимося на дерево гаплотипів. Дійсно, саме 15 мутацій. Порахуйте самі, це все «вилазять» з колеса спиці. А вилазять вони в різних (і заздалегідь непередбачуваних) місцях, тому що мутації відбуваються неупорядковано. 53 гаплотипа не мутувані, вони утворюють рівну гребінку, або «колесо» навколо центральної частини. Це - той же самий гаплотип, що мав спільний пращур всіх 68 осіб:
15 12 25 11 11 13
53 людини цей гаплотип повністю зберегли, тому що 625 років - це відносно малий час, і 15 мутацій - це все відхилення від пращурного гаплотипа, що за цей час змогло статися.
Показані вище шість чисел відповідають шестьом ділянкам в Y-хромосомі ДНК, в яких повторюються вибрані дослідниками короткі нуклеотидні послідовності. У першій ділянці - 15 повторів, у другому - 12 повторів, у третьому - 25 повторів, і так далі. Мутація - це зміна числа повторів. Помилився копіює фермент при біологічному копіюванні Y-хромосоми, скопіював першу ділянку не 15 разів, як заповідали пращури, а 16 раз, вийшло
16 12 25 11 11 13
Це - два ідентичних гаплотипа, на дереві вище під номерами 054 і 060.
З правого боку - гаплотип 061, він має вигляд
15 12 25 11 11 14
У нього мутація проскочила в останній ділянці, було 13 повторів, стало 14. Тобто знову фермент помилився в бік завищення числа повторів. А от у сімці ідентичних гаплотипів під номерами 055-057, 062, 064, 066, 068 - мутація пішла в третій ділянці, на пониження (25 → 24), статистика адже, і вийшло
15 12 24 11 11 13
Дійсно, ця ділянка, або маркер, як їх називають, один з найбільш схильних до мутацій. Теорія підказує, що в першій ділянці на всі гаплотипи - дві мутації, в останньому - одна мутація, в третьому - сім мутацій. Так і вийшло на практиці. Тобто навіть при такій малій статистиці закономірності в цілому виконуються. Більш того, всі інші п'ять мутацій на дереві гаплотипів відносяться до того ж третього швидкісному маркеру. Гаплотипи під номерами 059, 063 і 065 мають вигляд
15 12 26 11 11 13
тобто мутація пройшла на одиницю на підвищення, 25 → 26. Останні дві мутації, в гаплотипі під номерами 058 і 067, виявилися подвійними (25 → 23, 25 → 27):
15 12 23 11 11 13
15 12 27 11 11 13
Вони й сидять по обидві сторони «букета», тому що не походять одна від іншої. Зауважте, що довжина «спиці» у двох останніх випадках теж подвійна, порівняно з усіма іншими. Тобто по виду дерева гаплотипів можна сказати, які мутації поодинокі, які - подвійні або потрійні.
Все це відбувається тому, що так налаштована професійна комп'ютерна програма, яка будує дерево гаплотипів на підставі введеного списку гаплотипів. Вона сортує гаплотипи по гілках, тобто по схожості гаплотипів, сортує по тому, в яких маркерах відбулися мутації, скільки мутацій відбулося, і по тому, наскільки гілки древні. Тому по виду дерева людина досвідчена відразу може зрозуміти, яка структура тієї чи іншої популяції, скільки в ній основних гілок, який відносний вік гілок, і далі по числу мутацій в гілках розрахувати, коли жив загальний пращур кожної гілки.
По виду базових гаплотипів в різних регіонах і за віком їх гілок можна встановлювати, звідки, куди і коли проходили стародавні міграції людей і де зараз живуть їхні нащадки. Тобто можна проводити ДНК-генеалогічне картування регіонів, материків, і всієї планети, як у просторі, так і в часі. У цьому - методологія ДНК-генеалогії. А далі йдуть інтерпретації одержуваних даних в термінах історії, мовознавства, антропології, стиковка їх з відомими даними (тоді це по суті «калібрування»), або з даними слабкою, сумнівними, конфліктними - тоді це додаткова «точка опори», або, нарешті, це введення в науковий обіг зовсім нових даних, і відкриття шляху для їх верифікації, перевірки, обговорення, висунення нових гіпотез і положень.
Ось навіщо цей розділ у нарисі про ДНК-генеалогії кавказців. Без нього було б важко зрозуміти, навіщо наведені дерева гаплотипів, як на них дивитися, і що вони дають.
Наступний розділ ми почнемо з розгляду загального дерева гаплотипів західного і центрального Кавказу. Гаплотипи посередні, всього 19-маркерні (потім покажемо 37- і 67-маркерні), але і з них можна дещо витягнути, як буде видно. Але ми змушені виходити з того, які гаплотипи знаходяться в нашому розпорядженні. У цьому відношенні Кавказ вивчений далеко не повністю, але що можна зробити, якщо кавказці тільки починають тестувати свої гаплотипи? Поки самі кавказці не зрозуміють важливість цих досліджень, ніхто за них це завдання не вирішить.
Проте, багато чого вже зроблено, завдяки двом майже не перетинаючих джерел інформації - це ентузіасти з Кавказу, які посилають зразки своїх ДНК в комерційні компанії, платять за це гроші і виставляють протяжні (67- і 111- маркерні) гаплотипи і гаплогрупи в бази даних для наукових досліджень, і професіонали, як правило, популяційні генетики, які проводять тестування за вибірками за всіма правилами, але задовольняються в кращому випадку тільки 19-маркерними гаплотипами.
Продовження: Що говорить ДНК-генеалогія про кавказців - 2
Анатолій А. Кльосов,
професор, доктор хімічних наук
В тему: